Les arbres, des timides ?
La fameuse timidité des cimes, qui voudrait que les arbres soient « polis » et que leurs branches maintiennent un respectueux espace entre elles, s’avère plus subtile que cette simple vision de relations sociales sous un angle anthropomorphique. Explications.

En cette période où l’émotion et la sensibilité prêtées aux plantes émerveillent, interrogent, font débat, voire polémique, et amènent des réflexions sur les notions de neurobiologie végétale (Barlow, 2008), il existe un phénomène observé depuis longtemps par des forestiers au sujet de la concurrence des couronnes : les individus de certaines espèces, au cours de leur développement, montrent des couronnes qui s’interpénètrent tandis que d’autres s’évitent. Pour caractériser ces dernières, l’expression de « crown-shyness » (timidité des couronnes) a été proposée par Jacobs (1955). En 1977, Francis Ng, forestier malaisien, lors de la description de grands arbres asiatiques (Dryobalanops aromaticum, Dipterocarpaceae), montre qu’il y a aussi de la timidité intra-couronne (photo ci-dessous). Ce terme exprime une sensibilité de la plante par rapport à ses voisins ou entre branches. Mais qu’en est-il exactement et comment analyse-ton actuellement ce phénomène ?

La timidité des couronnes : une observation
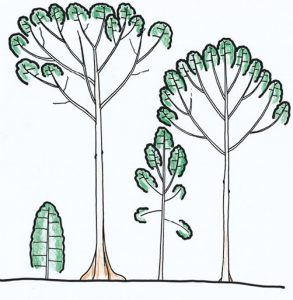
individualisation des cimettes chez Dryobalanops aromaticum
(d’après Hallé & Ng, 1981).
Lors du développement de l’arbre, la couronne de certaines espèces montre une délimitation de plus en plus marquée avec la couronne des individus voisins. Cela se traduit par un espace vide plus ou moins régulier tout autour de la couronne. Ce comportement s’observe chez des espèces de différents groupes taxonomiques et dans différentes formations végétales.
De nombreux travaux ont été consacrés à la modélisation de ce comportement chez le Pinus contorta avec des objectifs de production et de gestion forestière. Ils mettent en évidence, chez cette espèce, les paramètres de la croissance accompagnant cet isolement des couronnes individuelles. Des essences comme le Pinus pinea (pin pignon) montrent une interpénétration des couronnes dans les jeunes stades du développement puis, à l’état adulte, les couronnes ont une forte tendance à s’isoler.
Ce comportement peut s’observer entre couronnes d’individus différents mais aussi à l’intérieur d’une même couronne individualisant des sous-ensembles de la couronne parfois appelés des cimettes (Figure n° 2A). Cette timidité intra-couronne a été illustrée par Francis Hallé et Francis Ng sur les arbres de la famille des Dipterocarpacées (1981) en utilisant la notion de réitération pour préciser la mise en place de ce phénomène (Figure n° 2B). Cette notion, classique en architecture végétale, correspond au développement d’individus nouveaux au sein de la couronne d’un arbre existant.

La timidité des couronnes : des mécanismes
Divers mécanismes sont envisagés et envisageables pour expliquer la timidité des couronnes.
L’abrasion : Chez le chêne vert, après un coup de vent, de nombreux petits rameaux morts secs s’observent au sol. Chez le pin pignon, l’observation des extrémités des tiges sur le pourtour de la couronne montre des cassures, des frottements plus ou moins forts, des bourgeons râpés. Cette hypothèse simple a été démontrée sur des couronnes de grands palétuviers (Avicennia germinans). Plus récemment, Hajek et al. (2015) ont montré que ce phénomène d’abrasion mécanique était le mécanisme majeur des interactions dans les couronnes d’une forêt mixte tempéré (hêtre, charme, tilleul).
La lumière : L’ombrage porté par les couronnes voisines est un deuxième facteur qui a aussi été démontré en 1984, puis de manière répétée. Il suggère des mécanismes impliquant des composantes de la lumière (rapport rouge/infrarouge, bleue) modifiées par la présence d’une plante voisine et induisant la réorientation des feuilles et l’allongement des tiges pour permettre une optimisation de l’interception de la lumière par les parties de la plante subissant l’ombrage (Ballaré, 1999). Plus récemment, des mécanismes de régulation génétique mettant en jeu des protéines et des activations de gènes spécifiques ont été suggérés.
L’hypothèse chimique : Si des hypothèses concernant l’implication de composés chimiques ont été évoquées plus ou moins précisément, rien n’a vraiment été mis en évidence. Récemment, un auteur évoque l’existence de composés camphrés dans les feuilles de Dryobalanops aromaticum (Dipterocarpacées) et d’Eucalyptus spp (Myrtacées) et leurs effets inhibiteurs sur la croissance de plantes d’autres espèces. Il émet une hypothèse causale entre ces deux faits, rapprochant ces phénomènes de la timidité.
Si l’émission et le rôle de composés organiques volatils sont bien identifiés dans les mécanismes de défense contre des insectes phytophages, la causalité directe entre ces molécules et la compétition entre plantes n’est pas pour l’instant démontrée in situ, mis à part pour l’éthylène. Les effets de ce composé sont en effet comparables au syndrome d’évitement de l’ombrage (orientation foliaire et allongement des tiges ; Pierik et al., 2013).
La timidité des couronnes : une hypothèse intégratrice et des rôles qui commencent à être bien identifiés
Une réflexion récente passe en revue les hypothèses que nous venons de voir en concluant que la timidité des couronnes semble résulter d’une multiplicité de causes et que les différents écanismes sont liés entre eux. On peut envisager une suite de mécanismes imbriqués : évitement via la détection du voisinage par les phytochromes, renforcés par l’action éventuelle de composés organiques volatiles ; évidage via l’abrasion mécanique liée à la stratégie de réitération, aux mécanismes d’élagage et aux propriétés mécaniques du bois, chacun de ces mécanismes pouvant s’exprimer de manière plus ou moins intense selon l’espèce.
Les vides autour des couronnes (« canopy gaps », « canopy channels ») sont des espaces d’échange pour les organismes volants (animaux mais aussi graines). Ces vides assurent de la fluidité pour les échanges entre couronnes mais aussi pour les déplacements verticaux entre les différents niveaux de la forêt. À l’inverse, cela interdit des passages directs entre couronnes voisines (par exemple pour les fourmis). Ces passages deviennent possibles quand il y a des lianes qui assurent le relais, mais… plus il y a de vent, plus il y a d’abrasion, plus les couronnes sont timides et plus les lianes ont du mal à coloniser un arbre voisin…
Yves Caraglio
UMR-AMAP Botanique et modélisation de l’architecture des plantes et des végétations, département systèmes biologiques (Bios) du Cirad
À LIRE
Ballaré, C. L. Keeping up with the Neighbours: Phytochrome Sensing and other Signalling Mechanisms. Trends in plant science 1999;4(3):97-102
Hajek, P., Seidel, D., Leuschner, C. Mechanical Abrasion, and not Competition for Light, is the Dominant Canopy Interaction in a Temperate Mixed Forest. Forest Ecology and Management 2015;348:108-116
Hallé, F., Ng, F. S. P. Crown Construction in Mature Dipterocarp Trees. The Malaysian Forester 1981;44(2-3):222-233
Pierik, R., Mommer, L., & Voesenek, L. A. Molecular Mechanisms of Plant Competition: Neighbour Detection and Response Strategies. Functional Ecology 2013;27(4):841-853
Un lien sur un site qui bouge !
https://www.youtube.com/watch?v=EqFmLgK_H9s