Histoire et diversité des Asteraceae (ou Compositae)
Quel est le point commun entre une marguerite, une camomille, un bleuet, une chicorée ou… une endive ? Tous appartiennent à la grande famille des Asteraceae, dont voici l’histoire phylogénétique.

Figure n° 2 (en bas) : Les trois types d’inflorescences d’Asteraceae en fonction des types de fleurs présents. A gauche : Tubuliflore, au centre : Liguliflores, à droite : Radiées (modifié d’après Régnier V. & Chadefaud M. 1939 Histoire naturelle première année. Librairie Delagrave éd.).
La famille des Asteraceae (Astéracées en français), aussi appelée Compositae (Composées en français) est la famille de plantes à fleurs qui rassemble le plus grand nombre de genres (plus de 1 900) et d’espèces (plus de 33 000). En cela, elle dépasse largement les numéros 2 et 3 que sont les Orchidaceae (900 genres, 27 800 espèces) et les Fabaceae (750 genres, 19 500 espèces).
Les Astéracées de plus près
Au même titre que d’autres familles, les Astéracées font partie de ces groupes qui ont été reconnus bien avant l’avènement des classifications scientifiques, du fait de la présence d’inflorescences très particulières, les capitules. Ces capitules, qui sont des structures composées parfois de plusieurs centaines de minuscules fleurs, miment une fleur de très grande taille, et ce que l’on croit être plusieurs dizaines de sépales sont en fait des bractées. C’est un piège classique de l’apprentissage de la botanique, une fleur de Marguerite n’a pas d’une quinzaine à plus de trente pétales blancs et plus d’une centaine d’étamines mais bien une quinzaine à plus de trente fleurs ligulées, chacune ayant cinq pétales et cinq étamines, et une centaine de fleurs tubulées, elles aussi à cinq pétales et cinq étamines (Figure n° 1). D’un point de vue évolutif, ces inflorescences, mimant une très grosse fleur, sont particulièrement adaptées à la pollinisation par les insectes lécheurs-suceurs qui n’ont qu’à se poser sur ce « terrain » pour à la fois se nourrir et se mettre des quantités de pollen sur les pattes et le corps.
En plus de ces capitules, la famille des Asteraceae présente, de manière caractéristique, des fleurs à pétales soudés entre eux et à étamines soudées aux pétales par leur base et entre elles par leurs anthères. Autre caractéristique, les fruits sont des akènes issus d’un ovaire infère et surmontés des restes des sépales. Ceux-ci constituent un pappus plus ou moins développé qui assure la dissémination par le vent. La libération du pollen est aussi caractéristique de la famille : la croissance du style et des stigmates, passant au travers d’un cylindre formé par les anthères, libère le pollen qui se retrouve sur le dos du stigmate.
Organisation des capitules et classement
Classiquement, on reconnaît trois types d’organisation des capitules, qui correspondent plus ou moins à trois groupes dans les Asteraceae. Des espèces dont les capitules sont composés exclusivement de grandes fleurs tubulées forment le groupe appelé Tubuliflores ou Cynarocéphales (exemple : le bleuet), des espèces dont les capitules ne contiennent que des fleurs ligulées, constituent un groupe nommé Liguliflores ou Chicoracées (exemple : le pissenlit), et des espèces dont les capitules contiennent de petites fleurs tubulées au centre et des fleurs ligulées en périphérie, sont celles que l’on nomme les Radiées ou Corymbifères (exemples la marguerite ou la camomille) (Figure n° 2).
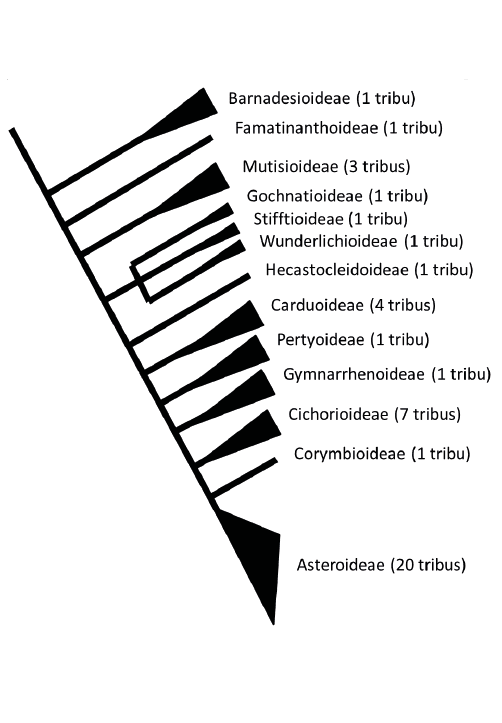
Aujourd’hui, cette distinction en trois ensembles sur la seule base de l’organisation de l’inflorescence ne tient plus, les études des années 1970-1980 et les données moléculaires du XXIe siècle ont sensiblement modifié la classification de la famille. Après une période, dans les années 1980, où divers systèmes existaient en parallèle (3 sous-familles et 13 tribus, 3 sous-familles et 17 tribus, 5 sous-familles et 30 tribus), la transcription de la phylogénie de la famille en classification conduit à reconnaître 13 sous-familles et 30 tribus (Figure n° 3 – d’après Funk et al. 2009, Panero et al. 2014, Fu et al. 2016).
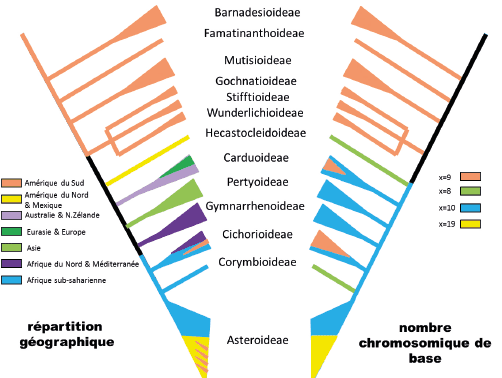
Du point de vue chromosomique, d’assez nombreuses modifications des chromosomes auraient eu lieu au cours de l’évolution de la famille. Le nombre de base primitif serait de 9 avec, dans les diverses sous-tribus basales, une forte tendance à des évènements de polyploïdie (doublement du nombre de chromosomes de base) ou de dysploïdie (modification du nombre chromosomique de base par fracture d’un chromosome en deux, ou par fusion de deux chromosomes en un seul). Une modification notable du nombre de base apparaît cependant à partir de la sous-famille des Cichorioideae, avec une modification du nombre de base passant de 9 à 10 chromosomes. Cette modification s’est accompagnée d’un changement de répartition géographique (ou de capacité de dispersion). Les lignées primitives de la famille (celles à x = 9) sont sud-américaines alors que les lignées dérivées (à x = 10) ont colonisé tous les continents.
De plus, au sein de la sous-famille des Asteroideae, le clade nommé « Heliantheae alliance » présente un nombre de base de x = 19. Cette modification pourrait correspondre soit à la résultante d’une hybridation d’un ancêtre à x = 10 avec un autre à x = 9, suivie d’une diploïdisation, soit à la diploïdisation d’un taxon tétraploïde (à x = 10 et 2n = 4x = 40) dont le caryotype (initialement à x = 20 et 2n = 2x = 40) se serait ensuite stabilisé par disploïdie (d’où x = 19 et 2n = 2x = 38) (Figure n° 4 à droite, d’après Semple et Watanabe 2009). Quelle que soit la complexité apparente de ces événements anciens, des modifications similaires ont été étudiées de très nombreuses fois dans la famille, à des échelles taxonomiques plus fines, au point que les Asteraceae sont un modèle pour l’évolution des chromosomes (en particulier le genre Tragopogon).
Une distribution géographique cohérente
Comme mentionné un peu plus haut, la distribution géographique des Asteraceae montre aussi une structuration cohérente avec sa phylogénie. Les premières lignées divergentes sont sud-américaines et, de manière concomitante à une modification d’organisation chromosomique, des lignées ont colonisé l’Amérique du Nord, l’Australie et la Nouvelle-Zélande, l’Asie, l’Afrique du Nord et la Méditerranée, ainsi que l’Afrique sub-saharienne. Dans la sous-famille des Carduoideae, une tribu, les Cardueae, a colonisé l’Eurasie et l’Europe ; dans la sous-famille des Cichoirioideae deux tribus (Gundelieae et Cichorieae) ont colonisé l’Afrique du Nord et la Méditerranée et une autre tribu, les Liabeae, a recolonisé l’Afrique du Sud. De plus, au sein des Asteroideae, l’essentiel du clade de l’« Heliantheae alliance », aurait recolonisé l’Amérique du Nord et le Mexique.
En plus de ces modifications à divers niveaux, les plantes dont l’homme fait un usage alimentaire sont principalement issues de quelques tribus, Cichorieae (ou lactuceae, sous-famille des Cichorioideae), Cardueae (ou Cynarieae, sous-famille des Carduoideae), Heliantheae et Anthemideae (sous-famille des Asteroideae) (pour plus de détails voir Chauvet, 2019). Ces tribus sont également celles fournissant un bon nombre des genres ornementaux de la famille. Plus qu’une explication phylogénétique, il faut plutôt associer cela aux aires de répartition de ces tribus, qui concordent avec celles d’un certain mode de société humaine.
Valéry Malécot
Maître de conférences en botanique à Agrocampus Ouest et chercheur au sein de l’UMR, IRHS (INRA / Agrocampus Ouest / Université d’Angers)
BIBLIOGRAPHIE
Chauvet M. 2019 Encyclopédie des plantes alimentaires. Belin, Paris.
Semple J.C. & Watanabe K. 2009 A Review of Chromosome Numbers in Asteraceae with Hypotheses on Chromosomal Base Number Evolution. Chapter 4 Pp.61-72 in Funk V.A., Susanna A., Stuessy T.F., Bayer R.J. (editors). Systematics, Evolution, and Biogeography of Compositae. International Association for Plant Taxonomy, Vienna.
Abeilles et astéracées : Production de miel assurée !

Les espèces de la famille des astéracées contribuent de manière particulièrement importante à la production de miel. Leur pollinisation est généralement assurée par des insectes car ils y trouvent facilement du nectar en raison de la disposition des organes floraux, regroupés et serrés en capitules.
Parmi le nombre considérable d’espèces de cette famille, les plus importantes pour la production de miel le sont par leur fréquence, en tant qu’espèce cultivée ou spontanée, par leur potentiel de production de nectar et (ou) de pollen ou, enfin, par la durée et (ou) la période de leur floraison.
Les principales astéracées mellifères
Parmi les espèces ornementales, il faut citer les échinops (Echinops spp.), le bleuet (Centaurea cyanus), les asters (Aster spp.), les rudbeckias, l’eupatoire chanvrine (Eupatorium cannabinum), le souci (Calendula officinalis), les cosmos (Cosmos spp.), la doronic du Caucase (Doronicum orientale), les gaillardes (Gaillardia spp.)… Toutefois, certaines espèces, telles que les dahlias, souvent sélectionnées par les horticulteurs pour d’autres caractéristiques, sont peu mellifères.
En grande culture, le tournesol (Helianthus annuus) est cultivé sur des surfaces importantes. La visite des capitules par les abeilles est fondamentale pour la production grainière. Chez les espèces spontanées, les pissenlits (Taraxacum spp.) sont généralement cités en premier car leur fréquence et leur précocité sont particulièrement bénéfiques, même si les jardiniers se désolent parfois de leur présence. De très nombreuses autres espèces sauvages participent à la production mellifère, par exemple la centaurée jacée (Centaurea jacea), différents chardons (Carduus, Cirsium…), l’achillée millefeuille (Achillea millefolium), la grande bardane (Arctium lappa), l’arnica (Arnica montana), la verge d’or (Solidago virgaurea), la tanaisie (Tanacetum vulgare), la grande camomille (Tanacetum partenium), l’inule visqueuse (Dittrichia viscosa)… N’hésitez pas à leur laisser une petite place dans vos jardins ! En revanche, dans les jardins potagers, les astéracées cultivées telles que les laitues et chicorées, les artichauts, les cardons et les salsifis, bien que mellifères, sont récoltées avant leur floraison et ne profitent donc pas aux abeilles.
Jean-Daniel Arnaud
Membre du conseil scientifique de la SNHF