De la pollinisation à la formation des graines et des fruits
Françoise Corbineau , Emmanuel Gendreau

… Le pollen va germer, puis les gamètes mâles seront acheminés jusqu’au gamète femelle situé dans le sac embryonnaire de l’ovule grâce au tube pollinique. La fusion du gamète mâle et du gamète femelle ou « fécondation » donnera naissance au zygote, futur embryon de la graine. La pollinisation est donc indispensable à la fécondation à l’origine des graines. Elle est aussi nécessaire à la production des fruits et joue un rôle important dans le rendement des cultures et la qualité de certains fruits (taille, forme, …).
De l’ovule à la graine : Rôle de la fécondation
L’ovule, gamète femelle.
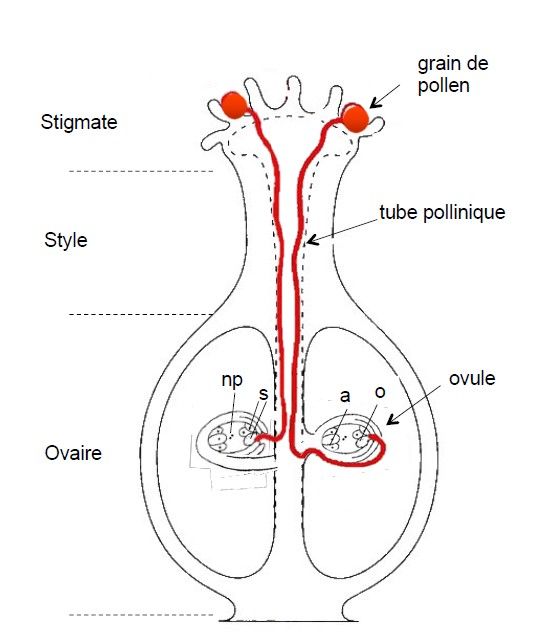
Chez les Angiospermes, l’ovule est généralement limité par deux téguments et présente un orifice, ou micropyle, à son extrémité. L’ovule est constitué d’un tissu homogène diploïde[1], le nucelle. Il est lié au carpelle au niveau du hile.
Dans le nucelle, une cellule proche du micropyle donne naissance à 4 cellules par méiose dont 3 avortent. La cellule restante, haploïde[2] (1n chromosomes), se divise pour former les 8 cellules du sac embryonnaire. L’oosphère (gamète femelle) se situe au niveau du micropyle, encadrée par 2 synergides. Les deux noyaux au centre du sac (noyaux polaires) fusionnent constituant ainsi un noyau secondaire diploïde, et 3 cellules antipodes restent au fond du sac embryonnaire (Figure 1).
On distingue 3 types principaux d’ovules chez les Angiospermes selon les positions respectives du hile et du micropyle: les ovules droits ou orthotropes pour lesquels, le hile et le micropyle sont opposés (Cistacées, Polygonacées, Juglandacées, Urticacées, Platanacées,…) ; les ovules campylotropes qui se courbent sur eux-mêmes (Cannabinacées, Caryophyllacées, Chénopodiacées, Crucifères, Papilionacées,…) et les ovules anatropes, la forme la plus courante, dont le micropyle se trouve proche du hile.
La germination du grain de pollen et la fécondation.
Une fois déposé sur le stigmate, le grain de pollen s’hydrate et produit un tube qui pénètre dans le style et progresse jusqu’au sac embryonnaire. L’acheminement jusqu’à l’oosphère est facilité par la proximité du hile dans le cas des ovules anatropes, alors que le tube pollinique doit s’engager dans la loge carpellaire pour atteindre le hile des ovules orthotropes (Figure 1). Le noyau végétatif localisé à l’extrémité du tube disparaît alors que le noyau reproducteur se divise en deux pour donner les deux gamètes mâles.
Les Angiospermes, tels que les fruitiers, sont caractérisés par une « double fécondation ». Un des gamètes mâles va fusionner avec l’oosphère pour donner un œuf diploïde qui se divise et donne l’embryon. La fusion du deuxième gamète avec les noyaux polaires donnera naissance à un tissu de réserves triploïde (1n chromosomes paternels + 2n chromosomes maternels), l’albumen. Dans des conditions favorables, la fécondation suit la pollinisation de quelques heures à plusieurs semaines (9 à 120 h chez les pommiers, pruniers et cerisiers, 4 sem chez Citrus trifoliata). Chez certains chênes, 1 an peut séparer pollinisation et fécondation.
La fécondation ne peut se réaliser que s’il n’y a pas d’incompatibilité entre le grain de pollen et l’organe femelle. Ce phénomène permet d’éviter les croisements inter-génériques et interspécifiques. Toutefois, il existe aussi des cas d’auto-incompatibilité qui réduisent les croisements entre les gamètes mâles et femelles d’un même individu[3].
La graine : un emboitement de structures aux identités génétiques différentes.
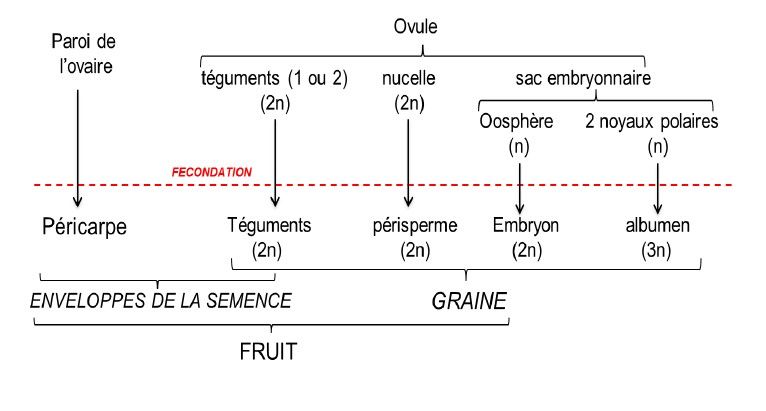
La figure 2 montre le devenir des différentes parties de l’ovule et de l’ovaire après la fécondation. La graine provient du développement de l’ovule : les téguments de l’ovule se transforment en téguments de la graine et sont diploïdes (2n, maternels) ; l’embryon diploïde (1n maternels + 1n paternels) se développe dans un tissu triploïde (2n maternels + 1n paternels), l’albumen, qui croit aux dépends d’un tissu diploïde, d’origine maternelle, le nucelle qui prend le nom de périsperme.
La pollinisation, facteur de production des semences.
La production des graines dépend évidemment de la pollinisation mais l’abondance du pollen intervient aussi, en particulier dans le cas des semences hybrides. Ainsi, dans le cas des semences de carotte, la distance des plantes mâle-stériles des rangs pollinisateurs, et donc le nombre de grains de pollen déposés, a une incidence majeure sur le nombre de semences produites, leur poids et leur qualité germinative. Il est donc généralement recommandé de mettre des pollinisateurs (mouches, abeilles …) pour améliorer la production et la qualité des semences.
Comment naissent les fruits
Le passage de la fleur au fruit ou « nouaison » est normalement consécutif à la fécondation. Si la nouaison a lieu sans fécondation, on parle de parthénocarpie et le fruit ne renferme pas de graines. En absence ou non de fécondation, la paroi de l’ovaire donne naissance aux tissus du fruit. Elle peut se développer soit en une structure succulente et aboutir ainsi aux fruits charnus comme les baies (raisin, banane, tomate …), les piridions (pomme et poire) et les drupes (olive, pêche, cerise …) ou bien évoluer vers une structure lignifiée à maturité dans le cas des fruits secs. Ceux-ci peuvent être déhiscents, ils s’ouvrent alors pour libérer les graines, comme par exemple les follicules (pivoine, magnolia …), les gousses (pois, fèves), les siliques (chou, radis, colza) ou indéhiscents comme les akènes (tournesol, renoncule), les caryopses (graminées) et les samares (érable, frêne).
Dans le cas de la sténospermocarpie, la pollinisation et la fécondation se déroulent normalement mais l’arrêt du développement de l’embryon entraine l’avortement des graines, ce qui aboutit à des raisins sans pépin[4].
Le développement de fruits parthénocarpiques est bien connu des horticulteurs et peut être recherché. Les plus classiques sont par exemple l’ananas, certains pamplemousses, les oranges Navel, les bananes communes, les clémentines, des pommes, et des poires[5].
Pollinisation et qualité des fruits

Le développement des fruits est contrôlé par la production d’auxine, hormone végétale, par les graines. Chez de nombreuses espèces de pommes et de poires il existe une corrélation positive entre la masse du fruit et le nombre de graines qu’il renferme. L’avortement des graines peut être responsable de la croissance irrégulière des fruits. La Figure 3 montre que la partie moins bombée de la pomme correspond aux loges ovariennes dont les graines ont avorté. Il est donc capital d’assurer une bonne pollinisation[6]. Le nombre de graines intervient aussi sur la couleur, le goût et la texture de la chair des Kakis et la teneur en sucres des poires.
Un rôle capital à protéger
La pollinisation est capitale pour les cultures car elle contrôle la production des graines et des fruits même si des exceptions existent. Une insuffisance de pollinisation entraîne des conséquences économiques graves car elle impacte les rendements horticoles et agricoles et la qualité des productions. Il est donc capital de protéger les pollinisateurs et de tenir compte lors de la plantation de la disposition des arbres pollinisateurs pour assurer une bonne pollinisation.
[1] 2n chromosomes d’origine maternelle
[2] La cellule proche du micropyle subit une méiose qui conduit à 4 cellules dont le nombre de chromosomes est réduit de 2n à 1n
[3] Voir dans ce dossier, l’encadré « Prévenir les autofécondations : L’incompatibilité chez les plantes supérieures », par Yvette Dattée
[4] Voir dans ce dossier « Du raisin sans insectes pollinisateurs », par Laurent Torregrosa
[5] Pour en savoir plus sur le sujet, nous vous invitons à lire l’article de Michel Pitrat dans le n° 633 de Jardin de France de Janvier-février 2015.
[6] Voir l’article de B. Vaissière et celui Y. Lespinasse dans ce dossier.