Des stratégies inhabituelles de pollinisation
Aline Raynal-Roques , Albert Roguenant


Si toutes les fleurs d’une inflorescence s’épanouissent en même temps, la production de pollen est massive, est-ce un avantage ? Par exemple, Helosis cayennensis, Balanophoracée sans chlorophylle des sous-bois d’Amazonie, vit aux dépens des arbres qu’elle parasite. Elle n’a ni racines, ni tiges, ni feuilles mais des suçoirs qui s’enfoncent dans le bois de son hôte et des inflorescences. L’inflorescence est couverte de bractées rougeâtres, en parapluies, sous lesquelles se développent des fleurs unisexuées.
A l’épanouissement, toutes les fleurs femelles d’une même inflorescence déploient simultanément leurs stigmates blancs (figure 1). Plus tard, les stigmates, fanés, seront tombés. Les fleurs mâles s’épanouiront alors toutes ensembles et produiront des gouttes de nectar.
Sur une même inflorescence, deux floraisons se succèdent donc : d’abord femelle, ensuite mâle. La disjonction dans le temps a pour conséquence que, dans une même inflorescence, les fleurs mâles ne peuvent pas polliniser les fleurs femelles. Le pollen doit être transporté par un insecte attiré par le nectar, vers les fleurs d’une autre plante, encore au stade femelle ; la fécondation ne peut intervenir qu’entre individus différents.
Décalage dans le temps
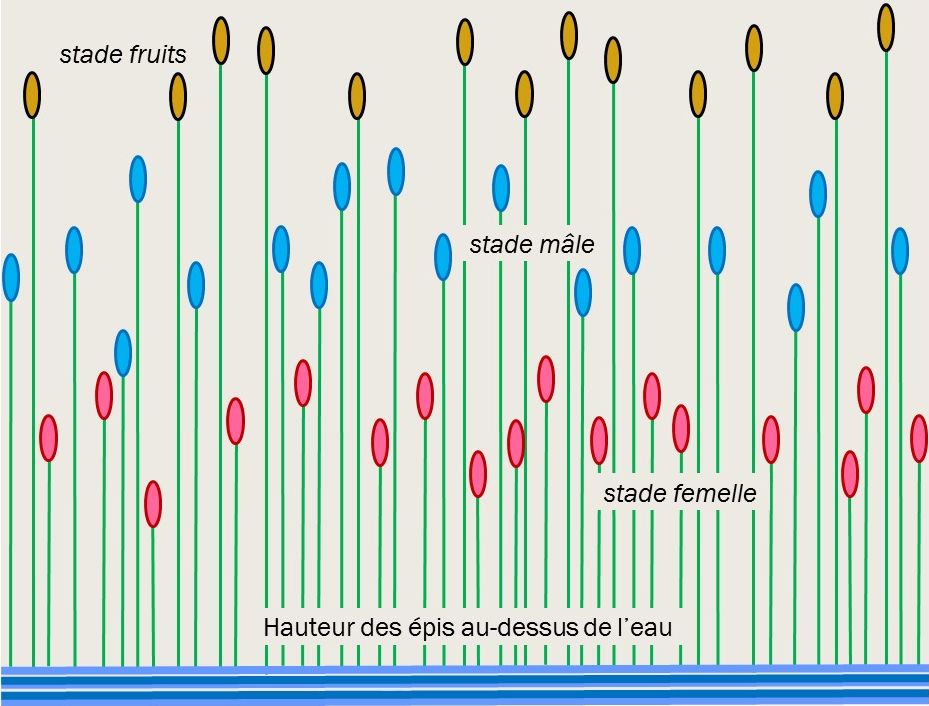
Un autre exemple, chez une Cypéracée des marais américains (Eleocharis interstincta).
J’ai pu observer une population de cet Eleocharis, dans les marais de Guyane. La tige nue porte un épi terminal de fleurs hermaphrodites et poursuit son allongement jusqu’à maturité de ses fruits.
Les fleurs d’un même épi s’épanouissent en deux temps. D’abord, les tiges sont hautes de 20 cm environ au-dessus de l’eau, toutes les fleurs de l’épi épanouissent leurs stigmates blancs, elles sont fonctionnellement femelles. Lorsque les tiges atteindront environ 35 cm, les stigmates seront fanés. Toutes les fleurs, alors fonctionnellement mâles, épanouiront leurs étamines (figure 2).
Le pollen libéré massivement en hauteur se dispersera sur les épis au stade femelle, situés en-dessous.
Dans une même inflorescence, la production massive de pollen d’une part, la maturité de tous ses stigmates d’autre part, sont décalées dans le temps. La pollinisation croisée en est favorisée et plus efficace.
Fleurs agressives : une stratégie de pollinisation inattendue
Imaginez une saga inattendue : une fleur attire un insecte pollinisateur. Lorsqu’il s’y pose, la fleur le capture. Il tente de s’échapper de sa prison, s’agite, se salit de pollen, puis la fleur relâche son étreinte : l’insecte, indemne, s’enfuit. Un peu plus loin, il rencontrera une autre fleur délicieusement parfumée, qui … le capturera à son tour. Pendant sa deuxième détention, un stigmate pickpocket pourra le délester de son pollen.
Deux exemples de plantes australiennes qui ont des organes capables de mouvements rapides, réversibles, reproductibles, en réponse à un stimulus : leurs fleurs peuvent emprisonner temporairement un pollinisateur potentiel.
Insecte capturé et libéré
La fleur de Stylidium graminifolium (Stylidiacées), étale les quatre lobes de sa corolle rose. A sa gorge se dressent de petites glandes odoriférantes. Un organe spécialisé, la colonne, rabattue sous le plan des lobes, porte à son sommet deux étamines réduites à leurs anthères et deux stigmates : les organes mâles et femelles sont très proches, mais distincts.
Les stigmates d’une fleur pourraient-ils recevoir le pollen de celle-ci ?
Non ! Chez cette espèce, étamines et stigmates ne sont pas fonctionnels en même temps …
Lorsqu’une fleur s’épanouit, ses deux étamines, mûres, libèrent leur pollen. Ses stigmates, peu développés, sont immatures : la fleur est fonctionnellement mâle. Dans cette même fleur, deux jours plus tard, les deux étamines seront flétries et ne contiendront plus de pollen. Les deux stigmates, développés, seront gonflés et fonctionnels : la fleur sera devenue femelle.
Attiré par les glandes, un insecte se pose sur la corolle. Ce léger contact déclenche un mouvement si rapide de la colonne, qu’il n’a pas le temps de s’envoler. La colonne se redresse, pivote, s’abat sur son dos, sans le blesser. Il est maintenu quelques minutes, étamines et/ou stigmates appliqués sur son dos (figue 3). Pendant ce temps, soit le pollen tombe sur son dos, soit les stigmates s’y sont posés et collectent du pollen.
La colonne se relèvera peu à peu, l’insecte sera libéré. Il se posera sur une autre fleur qui le capturera à son tour. La colonne pourra réitérer son mouvement aussi longtemps que la fleur restera épanouie.
La fleur tête de canard
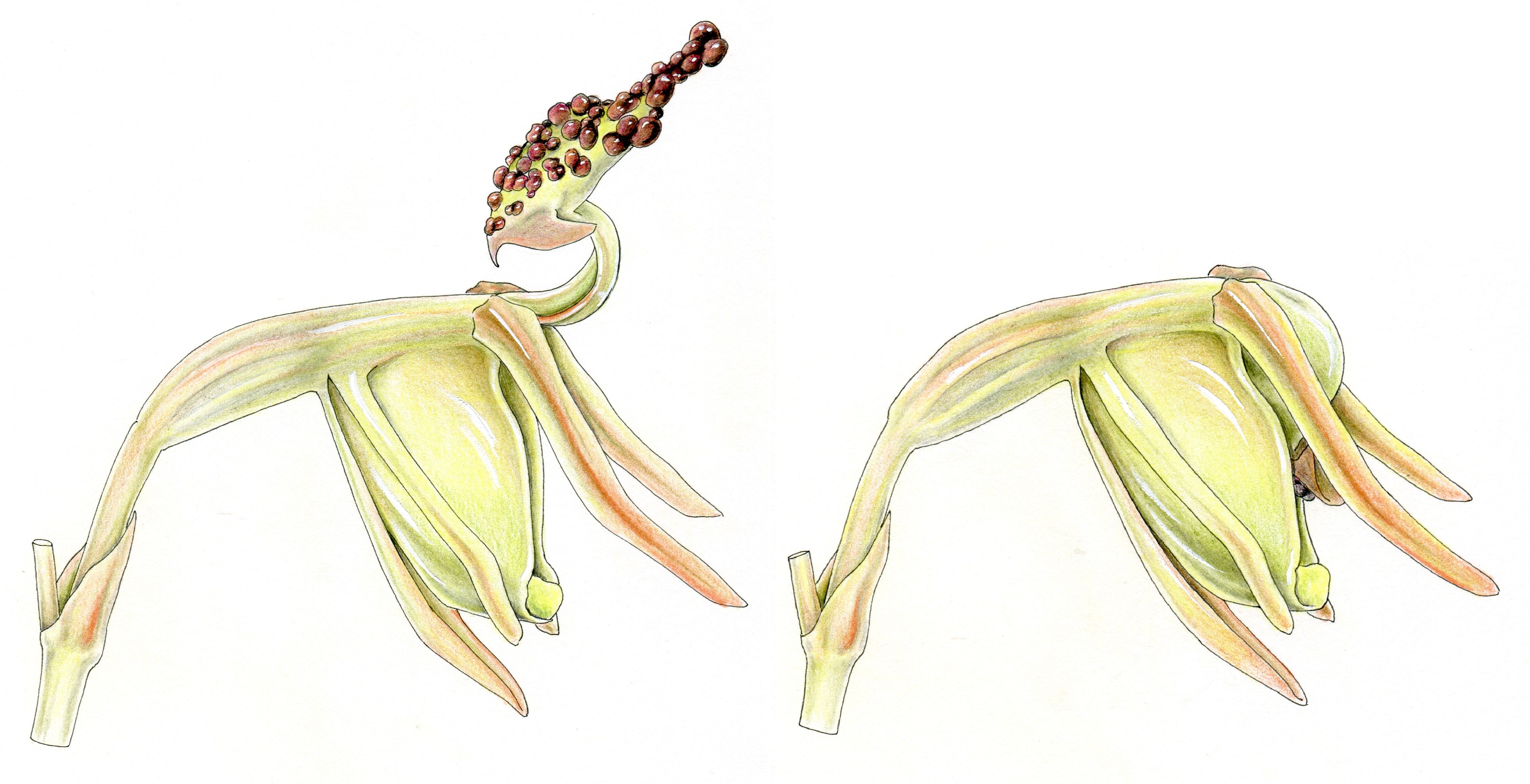
La fleur d’une petite Orchidée australienne, Paracaleana minor, évoque un canard en vol. La fleur semble seulement constituée d’une coupe verte et translucide. Elle est surmontée d’une lame étroite et élastique qui se prolonge en forme de « tête de canard ». La coupe contient le pollen, assemblé en pollinies, à son sommet, le stigmate est vers sa base. La « tête de canard » est couverte de glandes émettrices d’odeurs.
Lorsqu’un insecte se pose au milieu de la « tête », il active un point sensible et, en moins de 1/10e de seconde la « tête » s’incline, le « cou » inverse sa courbure, la tête s’encastre dans la coupe translucide (figure 4) … L’insecte enfermé se débat et abandonne du pollen sur le stigmate. Quelques minutes plus tard, le piège se rouvre lentement, l’insecte s’échappe, se frotte aux pollinies et les emporte vers une autre fleur de la même espèce.
Fleurs fermées et pollinisation.
Certaines fleurs ne s’épanouissent jamais, on les dit cléistogames, « mariage enfermé » ; elles restent à l’état de boutons dans lesquels s’effectuera une autopollinisation sans intervention extérieure, suivie d’une fécondation.
Les fruits résultant d’une cléistogamie produisent une descendance nombreuse, mais peu diversifiée.
La cléistogamie intervient souvent en réponse à une contrainte écologique. Dans un marais, une même plante peut porter des fleurs inondées cléistogames et des fleurs exondées épanouies ; si le niveau de l’eau baisse, les fleurs nouvellement apparues, s’épanouiront au-dessus de l’eau.
La nécessité de la rencontre d’un élément mâle et d’un élément femelle a conduit la nature à développer des stratégies parfois complexes et des outils, qui sont responsables du succès des végétaux de notre planète.
A quoi sert donc le pollen ?
La plupart des plantes à fleurs sont hermaphrodites. Elles produisent du pollen et des ovules, dans une même fleur ou dans des fleurs distinctes, sur la même plante.
Un grain de pollen est un individu vivant qui produira les spermatozoïdes. Ils féconderont l’ovule qui deviendra graine.
La pollinisation résulte de deux mécanismes :
– soit pollen et ovule proviennent d’une seule et même plante. Ils sont peu différents génétiquement et la descendance sera peu diversifiée mais nombreuse ;
– soit ils proviennent de deux plantes de la même espèce. Ils diffèrent génétiquement et la descendance, diversifiée, pourra développer des capacités d’adaptation et de résistance.
La pollinisation est la fonction essentielle qui engage l’avenir évolutif de l’espèce.
Pollen et rhume des foins
Les grains de pollens sont d’une grande diversité de formes et surtout d’ornementation.
Pourtant, le spécialiste de la santé ne les classera qu’en deux catégories : les pollens de grande taille, généralement véhiculés d’ailleurs par des auxiliaires justement nommés pollinisateurs, et les pollens de petite taille, confiés au vent pour rejoindre une fleur ou une inflorescence femelle. La dimension de ces derniers leur permet de s’insinuer passivement au plus profond de nos poumons, pouvant activer des réactions d’allergie plus ou moins graves, selon les individus.
Les conditions environnementales sont prédisposantes ou aggravantes, selon les cas.
Les allergologues et les pneumatologues sont aidés dans leurs diagnostics, mais aussi dans la prévention, par l’élaboration de calendriers polliniques et par les alertes émises par un réseau national de surveillance et de comptage des pollens contenus dans l’air.
Deux grandes périodes sont redoutables : le printemps, avec la floraison des pins et des arbres à chatons (saules, bouleaux, chênes…) et l’été, avec la floraison des prairies de graminées. Chacun connaît de réputation le rhume des foins !
Loin de méconnaitre leur importance, Jardins de France traitera bientôt des sensibilités polliniques dans le cadre d’un dossier consacré plus largement aux allergies imputables aux plantes.
Daniel Lejeune